Introduction
The study and understanding of the invertebrate eye is no less labor-intensive than that of the vertebrate eye, but it is further complicated by the size of the various components and by our general ignorance of the degree to which sensory information is processed within such tiny nervous systems.
As to the eye proper, we might begin by emphasizing—as in the vertebrate eye—its role as the collector and mediator of electromagnetic radiation emitted by the Sun. Such energy, in the form of light, may enter the eye directly when the invertebrate subject is oriented towards the Sun, or indirectly, as reflected light, when the subject is oriented towards objects within its visual sphere.
So what is the simplest information that might be acquired from light, requiring, in turn, the simplest amount of processing (which is of the essence in creatures with "nervous systems" containing only imaginable numbers of neurons)?
Initially, the subject may require little more than the knowledge of where light is or is not so that it may withdraw from sunshine. This would entail a rather small population of sensory cells whose "light"/"no light" signals could be used for simple negative phototaxis (i.e., movement away from the light stimulus).
For small organisms with short life spans in relatively small environmental niches, there may be no great evolutionary pressure to evolve visual sense organs of any greater sophistication.
However, it may have been the case that, where organisms were living in niches that had high levels of competition adversely affecting species survivorship, certain mutant progeny were experiencing greater success in predation. As a consequence, they lived relatively longer, healthier lives that resulted in higher levels of reproduction and, therefore, in the generalization of that mutant trait—now become a species-wide characteristic.
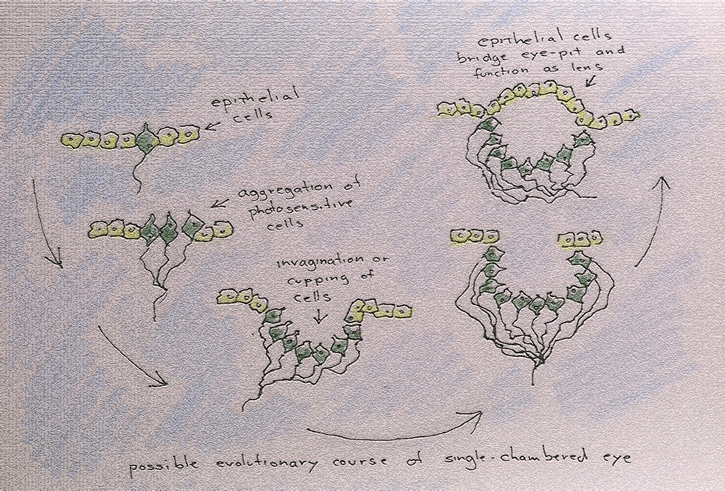
But what might this mutant trait have been? Well, photosensitive cells may have multiplied and arrayed themselves in such a fashion that no two such cells received light from the same direction (which would have resulted, for example, from the cupping of the cell layer in which the cells were embedded. This could have produced a concave shape as associated with the retinal layer of most vertebrate eyes or a convex shape as associated the compound eyes of such invertebrates as flies, mosquitoes and gnats). If this development were accompanied by a nervous system capable of comparing information from various cells, then the organism would have acquired directional vision. But this also necessitates the presence of a pigmented shield of some sort, a shield that would prevent light from scattering randomly throughout the eye and that would establish a difference between the front where light is streaming in and the back where light is being absorbed.
Development
Although molecular studies have resulted in revisions of previous assumptions regarding the multiple, but convergent, evolution of vertebrate and invertebrate eyes, it is still believed that the earliest "eyes" were collections of photosensitive cells gathered in pigmented depressions, such as the eyespots of planaria:
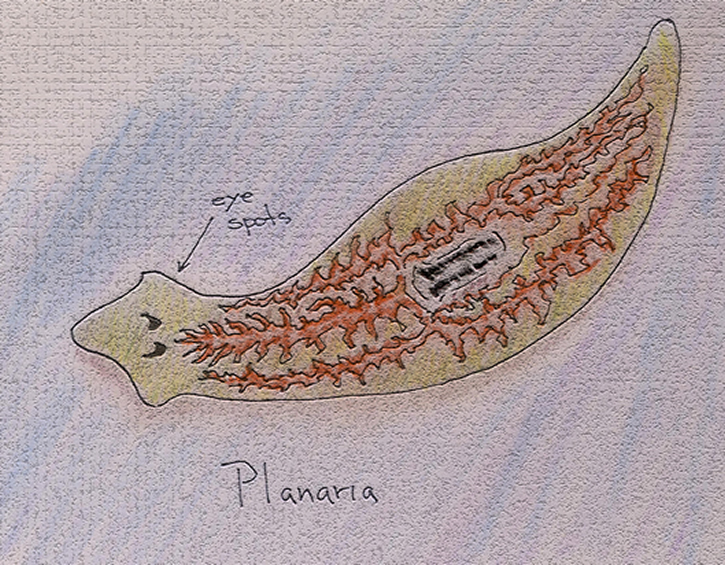
Cellular arrangements of this type can provide for some apparent sense of direction, which would enhance tactic behaviors involved in eluding moisture-robbing sunlight or predator-preceding shadows.
[image]
Camera-type eyes, found in both vertebrates and invertebrates, have a single chamber on one side of which is a lens that focuses an image on the photosensitive cells, or retina, located on the opposite side of the eye. In invertebrates, these types of eyes are referred to as ocelli.
Compound eyes, on the other hand, are common only to invertebrates. They are composed of an array of individual systems, ommatidia, each system having its own imaging apparatus, or lens, and its own collection of photoreceptors.
[Our apologies as this section is still to be edited]